Microtúbulo
Os microtúbulos são estruturas tubulares presentes nas células, de 25 nm de diâmetro exterior e cerca de 12 nm de diâmetro interior, com comprimentos que variam entre uns poucos nanómetros a micrómetros, que se estendem ao longo de todo o citoplasma. Encontram-se nas células eucarióticas e são formados pela polimerização de um dímero constituído por duas proteínas globulares, a α e a β tubulinas. Os microtúbulos nucleiam-se e organizam-se nos centros organizadores de microtúbulos (COMTs), como o centrossomo ou os corpos basais dos cílios e flagelos. Estes COMTs podem possuir centríolos ou não.
Os microtúbulos são heteropolímeros de α- e β-tubulina, as quais formam dímeros, que são a sua unidade estrutural. Os dímeros polimerizam em 13 protofilamentos, que logo se agregam lateralmente para formar estruturas cilíndricas ocas. Para polimerizar requer-se a presença de dímeros com uma concentração mínima determinada denominada concentração crítica, embora o processo se acelere pela adição de núcleos, que são alongados. Uma importante característica dos microtúbulos é a sua polaridade. A tubulina polimeriza por adição de dímeros num ou em ambos os extremos do microtúbulo. A adição faz-se por união cabeça com cauda durante a formação dos protofilamentos. Assim, vão-se formando fileiras de monómeros de α e β-tubulina na parede do tubo, o que provoca uma polaridade global ao microtúbulo. Devido a que todos os protofilamentos de um microtúbulo têm a mesma orientação, um extremo está composto por um anel de α-tubulina (denominado extremo -) e, o oposto, por um anel de β-tubulina (denominado extremo +).
Citoesqueleto
Como componentes do citoesqueleto (junto com a actina e os filamentos intermediários), dão à célula um suporte estrutural. Podem crescer e minguar para originar força.
Mobilidade de cílios
Os microtúbulos do axonema dos cílios e flagelos eucarióticos (os procarióticos são diferentes) permitem a mobilidade destes em associação com certas proteínas.
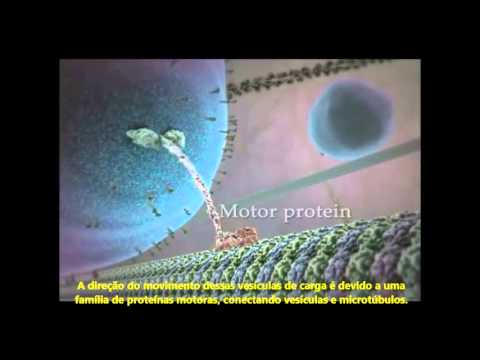
Transporte
Funcionam como autoestradas pelas quais se movem as proteínas motoras que arrastam vesículas, organelos e outros fatores celulares ao longo do microtúbulo. Estas proteínas aproveitam a hidrólise do ATP para gerar energia mecânica e mover-se sobre os microtúbulos. São a dineína, transportador retrógrado, e a quinesina, transportador anterógrado. A dineína é uma molécula de estrutura similar à quinesina: consta de duas cadeias pesadas idênticas que conformam duas cabeças globulares e de um número variável de cadeias intermediárias e de cadeias leves. Transportam desde o extremo (+) para o (-) do canal intramicrotubular. Pensa-se que a atividade de hidrólise de ATP, fonte de energia da célula, encontra-se nas cabeças globulares. A dineína transporta vesículas e organelos, pelo que deve interagir com as suas membranas, e, para interagir com elas, requer um complexo proteico, cujo elemento mais notável é a dinactina.
Mitose
Uma importante estrutura feita de microtúbulos é o fuso mitótico usado nas células eucarióticas para separar os cromossomos durante a divisão celular. A mitose é facilitada por um subgrupo de microtúbulos denominado microtúbulos astrais, definidos como microtúbulos originados no centrossomo que não se conectam com os cinetocoros dos cromossomos. Os microtúbulos astrais interagem com o córtex celular e ajudam na orientação do fuso. Colocam-se em disposição radial ao redor dos centrossomos. O grau de renovação ou reciclagem desta população de microtúbulos é maior que o de qualquer outra população. Os microtúbulos astrais funcionam concertadamente com as proteínas motoras dineínas, as quais estão orientadas com as suas cadeias leves unidas à membrana celular e a sua porção dinâmica unida ao microtúbulo. Isto permite que a contração da dineína empurre o centrossomo para a membrana, facilitando a citocinese.
No desenvolvimento
O citoesqueleto de microtúbulos é essencial durante os processos morfoxenéticos do desenvolvimento dos organismos. Por exemplo, durante a embriogénese na mosca da fruta Drosophila melanogaster, requer-se uma rede de microtúbulos intacta e polarizada dentro do ovócito a fim de estabelecer os eixos do ovo; deste modo, os sinais entre as células foliculares e as do ovócito (como os fatores semelhantes ao TGF-alfa) provocam a reorganização dos microtúbulos situando o seu extremo (-) na zona anterior do ovócito, o que polariza a estrutura e comporta o aparecimento de um eixo dorsoanterior. Este envolvimento na arquitetura corporal também se dá em mamíferos.
Regulação da expressão genica
O citoesqueleto celular é um elemento dinâmico que atua a muitos níveis na célula: além de dotá-la de uma forma determinada e de vertebrar o trânsito de vesículas e organelos, pode influir na expressão gênica. Porém, as vias celulares (ou seja, os mecanismos de transdução de sinal) que intervêm nesta comunicação são muito pouco conhecidos. Não obstante, descreveu-se a relação entre a despolimerização de microtúbulos mediada por fármacos e a expressão específica de factores de transcrição e, em consequência, a expressão diferencial dos genes dependentes da presença destes fatores. Esta comunicação entre o citoesqueleto e a regulação da resposta celular está também relacionada com a geração de fatores de crescimento: por exemplo, esta relação existe para o fator de crescimento do tecido conjuntivo.
A polimerização dos microtúbulos realiza-se num centro organizador de microtúbulos. Nestes centros existe um tipo de tubulina, chamada γ-tubulina, que atua nucleando a adição de novos dímeros, com intervenção de outras proteínas reguladoras. Considera-se que existe um complexo anular de γ-tubulina, sempre situado no extremo - do microtúbulo.
Instabilidade dinâmica
Durante a polimerização, ambas as unidades de tubulina estão unidas a uma molécula de guanosina trifosfato ou GTP. O GTP desempenha uma função estrutural na α-tubulina, mas é hidrolizado a GDP na β-tubulina. Esta hidrólise modula a adição de novos dímeros. Assim, o GTP hidroliza-se após um certo período de tempo, o que permite que, se a adição de dímeros é rápida, se forme no extremo (+) um casquete de β-tubulina unida a GTP, enquanto que, em caso de ser lenta, o que se expõe é tubulina unida a GDP. Esta união a um ou outro nucleótido é a que determina a velocidade de polimerização ou despolimerização do microtúbulo. Deste modo, um casquete no extremo (+) com GTP favorece a elongação, enquanto que um de GDP, a despolimerização.
Propriedades da polimerização da tubulina
Estas características explicam que exista uma inestabilidade dinâmica nos microtúbulos, que consiste em que, numa mesma célula, alguns microtúbulos estão despolimerizando-se (o que se chama catástrofe) e outros alongando-se (o que se chama resgate).
Alteração farmacológica da dinâmica microtubular
Existem uma grande quantidade de drogas capazes de unir-se à tubulina, modular o seu estado de ativação e assim interferir com a dinâmica microtubular a concentrações intracelulares muito mais baixas que a de tubulina. Deste modo, as células detêm o seu ciclo celular, o que pode conduzir à morte celular programada. Os compostos que modulam a atividade da tubulina podem dividir-se de forma geral em dois grandes grupos: em primeiro lugar estão os inibidores da sua polimerização, como a colchicina e a vincristina, que se unem a esta impedindo que forme microtúbulos. Por outro lado, estão os agentes estabilizantes de microtúbulos (MSAs), como o paclitaxel (conhecido comercialmente como taxol) e o docetaxel, os quais se unem preferentemente à tubulina montada, minimizando a dissociação da tubulina-GDP dos extremos dos microtúbulos e induzindo a montagem da tubulina-GDP normalmente inativa.